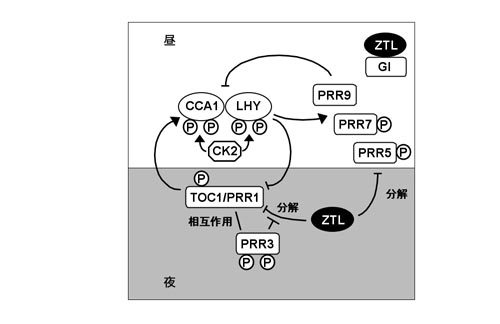
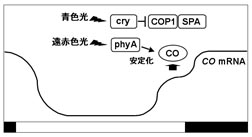
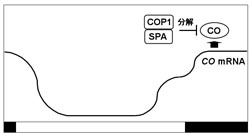
植物では、葉の就眠運動や花弁の開閉等、我々が肉眼で観察することが出来る現象だけではなく、様々な遺伝子の発現制御にも概日時計が関わっていることが知られている。シロイヌナズナの概日リズムは、おおまかに言えば、概日時計の中心振動体であるCCA1, LHYとTOC1のフィードバック制御を中心にして発生すると考えられている。MYB様転写制御因子をコードしているCCA1, LHY遺伝子は、TOC1によって発現が促進され、そのmRNA量は明け方に最大となる。他方、TOC1はプロモーター領域にCCA1, LHYが結合することで発現が抑制され、そのmRNA量が夕方から夜に最大に達する。TOC1はDNA結合領域を持たないことから、他の転写因子と相互作用してCCA1, LHY遺伝子の転写を正に制御すると考えられている。TOC1はPRRファミリーの一つで、PRR1とも呼ばれる。PRRファミリーの他の因子PRR3、PRR5、PRR7、PRR9も概日リズムの制御において中心振動体に近い役割を担っていると考えられている(図2)。
LKPファミリーの1つであるZTLの機能欠損点変異体(ztl-1, 2)では、標的タンパク質の認識に機能するkelchリピート上の保存されたアミノ酸に置換があり、概日時計の出力側であるCAB2遺伝子(早朝にmRNA量が最大となるmorning gene)の発現周期が3、4時間程度長くなり、CCR2遺伝子(夕方にmRNA量が最大となるevening gene)の発現周期や子葉の運動も長周期に変化している。機能欠損変異体とは対照的に、ZTLを過剰発現させた形質転換体ではその発現量に応じてCAB2プロモーター制御下にあるLUCレポーター遺伝子の発現周期が短くなったことから、ZTLは量依存的に概日時計を制御していると考えられている。
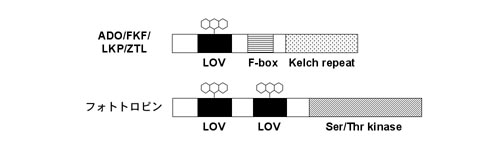
ZTLのLOVドメイン(LOV)は、光受容能を持つ点で非常にユニークである。LOVは、タンパク質間相互作用に関わるPAS (Per-ARNT-Sim)ドメインのサブセットの1つで、特に光,酸素,電位の変化に関与するものに共通した領域である。シロイヌナズナでは、LKPファミリーの他に、光屈性の青色光受容体であるフォトトロピン(phot)がLOVを有している(図1)。LOVにはFMN (flavin mononucleotide)等のフラビンが結合し、青色光照射によりフラビンとLOV内で保存されたシステイン残基との共有結合が誘起され,暗黒下で解離する(暗反転)。photではこのLOVの光反応により,タンパク質キナーゼ活性が制御されることが示されている。LKPファミリーのLOVはphotのLOVには無い短いループを持ち,光生化学的な性質が異なることが示されており、FKF1のLOVはアンチパラレルな方向で結合することで2量体形成に関与することが示唆されている。
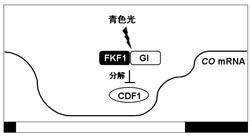
近年、ZTLのLOVドメインと巨大な核タンパク質であるGIが青色光依存的に相互作用することが報告された(図2)。ZTLのmRNA量は概日時計による制御を受けず一定だが、タンパク質量は夕方にピークを示す概日リズムを示す。このZTLの蓄積リズムにはGIの関与が示唆されている。ZTLとGIは相互作用すると互いに安定化し、giではZTLの夕方の蓄積量の増加が認められない。
TOC1及びPRR5タンパク質の蓄積量にもリズムが認められるが,このTOC1及びPRR5の分解はZTLが担っている (図2)。TOC1及びPRR5とZTLは酵母2-hybrid(Y2H)、免疫共沈降により相互作用が示され、ZTLのLOVドメインがTOC1及びPRR5との結合領域である。しかし現在までZTLとTOC1との結合に青色光による影響は認められていない。野生型では、TOC1タンパク質は夕方から夜にかけて、PRR5タンパク質は昼に蓄積するが、ztl変異体では常に高いレベルで蓄積する。野生型の細胞抽出液を用いたin vitro実験でTOC1及びPRR5タンパク質は速やかに分解されるが、ztl抽出液では分解されない。そして、この分解はプロテアソーム阻害剤によって阻害される。ZTLによるTOC1及びPRR5タンパク質の分解は暗黒下において顕著に促進する。ZTLとTOC1の相互作用はPRR3が競合的にTOC1と相互作用することで阻害される (図2)。PRR3タンパク質はTOC1タンパク質とほぼ同時期に蓄積しており、TOC1の安定化に寄与していると考えられている。ZTLとGIが光依存的に相互作用することでZTLの蓄積リズムが確立され、ZTLによるTOC1及びPRR5の分解によってTOC1及びPRR5の蓄積リズムの振幅が高まることで、結果として概日時計の周期と位相が正常に保たれると考えられている。
LKP2もY2HによりTOC1及びPRR5との相互作用が示されている。また、免疫共沈降法によりGIとの相互作用が示されている。LKP2の過剰発現体では連続明、連続暗下でのCAB2、CCR2遺伝子の発現と子葉の運動の概日リズムが消失し、長日条件下で花芽形成が著しく遅延する。これらはZTL過剰発現体の形質と非常に良く似ている。これらのことからLKP2は概日時計制御に関わっていると考えられる。 |